Central auditory pathway¶
Introduction¶
There is a collection of tracts that connects the cochlea to primary auditory cortex, known as the central auditory pathway. It passes through the major subcortical areas, which are characterized by groups of neurons known as nuclei, the plural of nucleus. Since I have given short shrift to the subcortical organs, the diagram from Chapter 3 is repeated here:
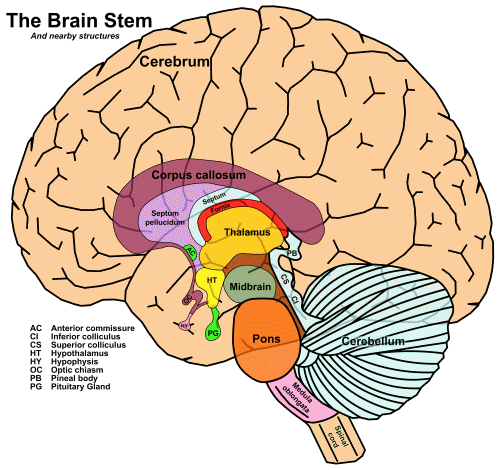
Fig. 52 The vertical axis of the brain [1]
The four major structures, from bottom to top, are the medulla oblongata, the pons, the midbrain – which make up the brainstem – and the thalamus.
The following drawing is the canoncial representation of information flow through this system:
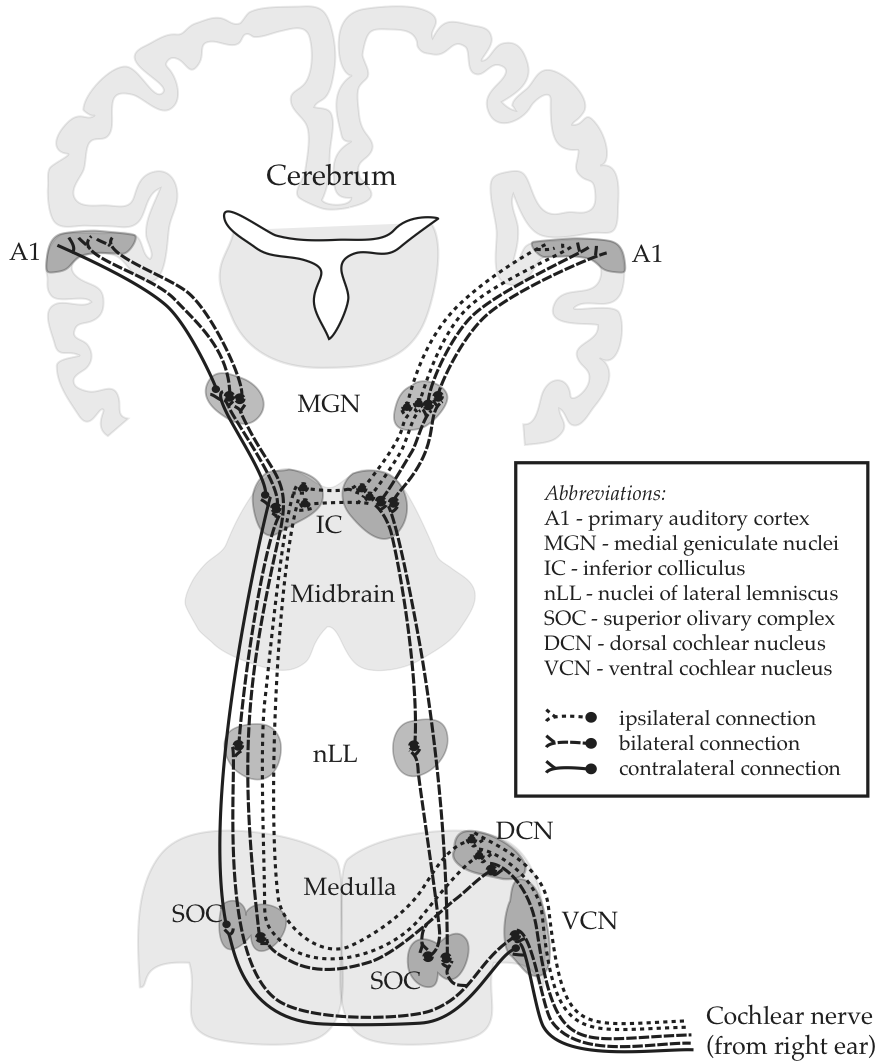
Fig. 53 The central auditory pathway [2]
It is highly significant that the drawing is labelled as Afferent. “Afferent” means ‘inward’ or ‘towards a central area’. In our particular case, it can mean ‘from sensory organs towards the brain’. A synonym that sounds more like normal English is ascending. By the way, the antonyms are efferent and ‘descending’, as ilustrated in the sidebar for a sensorimotor circuit.
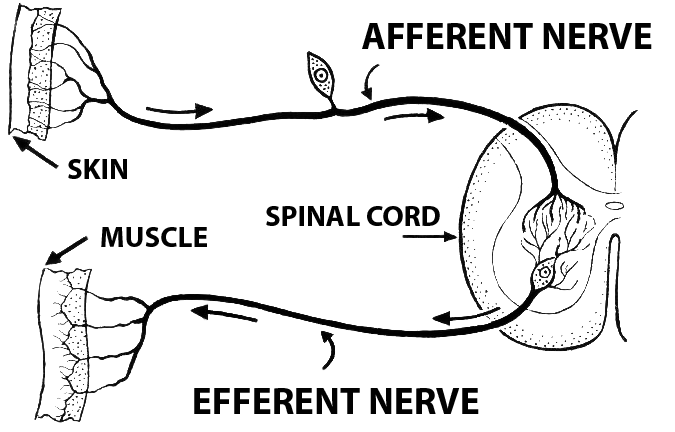
Stating information flow in this pathway as exclusively ascending begs the question of whether realist or constructivist perception is correct, since no rooom is made for potential descending information. We will come back to this problem again and again.
The brainstem¶
The cochlear nucleus¶
Dorsal cochlear nucleus¶
Tzounopoulos, Thanos. “Mechanisms of Synaptic Plasticity in the Dorsal Cochlear Nucleus: Plasticity-Induced Changes That Could Underlie Tinnitus.” textit{American Journal of Audiology} 17.2 (2008): S170-75.
The DCN is an auditory brainstem nucleus involved at the earliest stages of sensory processing and is believed to play a key role in orientation toward sounds of interest and/or suppression of responses to expected body-generated sounds (Shore & Zhou, 2006; Sutherland, Glendenning & Masterton, 1998; Young & Davis, 2001). DCN fusiform neurons receive input from the auditory nerve and from a system of parallel fibers carrying multimodal input, including proprioceptive signals encoding the position of the ears, as well as input from the vocal tract perhaps signaling suppression of body-generated sounds or vocal feedback (see Figure 1). DCN parallel fibers also activate inhibitory glycinergic cartwheel cells, which provide feed-forward inhibition to fusiform cells. The complex response of fusiform cells to auditory signals is shaped by feed-forward inhibition from additional cell types, the vertical cells of the deep layer (see Figure 1) and D-stellate cells of the ventral cochlear nucleus (Young & Davis, 2001).
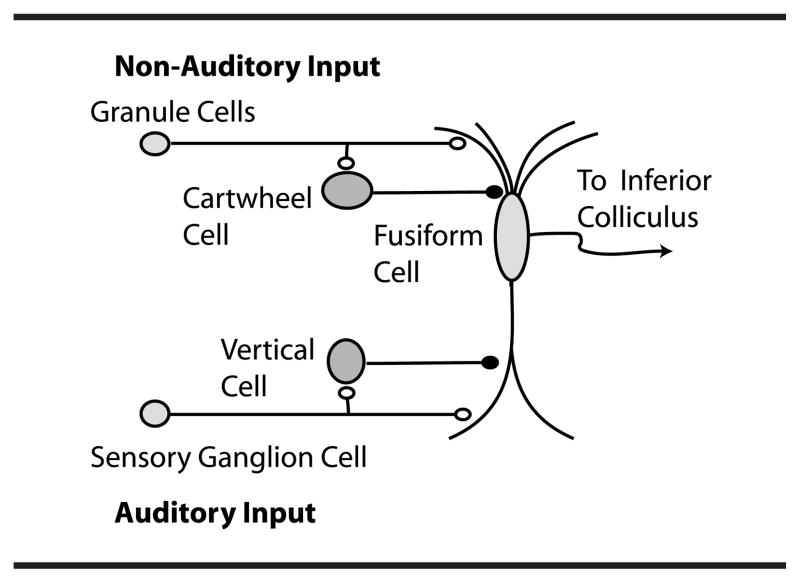
Fig. 54 Dorsal cochlear nucleus (DCN) circuitry
Ventral cochlear nucleus¶
Superior olivary complex¶
{Felix and Magnusson, 2016, neuro}
The superior paraolivary nucleus (SPON) is a prominent structure in the mammalian auditory brainstem with a proposed role in encoding transient broadband sounds such as vocalized utterances. … we measured evoked excitatory postsynaptic currents (EPSCs) in the SPON originating from the intermediate acoustic stria and compared them with the properties of EPSCs in the lateral superior olive (LSO) originating from the ventral acoustic stria during auditory development from postnatal day 5 to 22 in mice.
Before hearing onset, EPSCs in the SPON and LSO are very similar in size and kinetics.
After the onset of hearing, SPON excitation is refined to extremely few (2:1) fibers, with each strengthened by an increase in release probability, yielding fast and strong EPSCs. LSO excitation is recruited from more fibers (5:1), resulting in strong EPSCs with a comparatively broader stimulus-response range after hearing onset. Evoked SPON excitation is comparatively weaker than evoked LSO excitation, likely due to a larger fraction of postsynaptic GluR2-containing Ca2+-impermeable AMPA receptors after hearing onset. Taken together, SPON excitation develops synaptic properties that are suited for transmitting single events with high temporal reliability and the strong, dynamic LSO excitation is compatible with high rate-level sensitivity. Thus, the excitatory input pathways to the SPON and LSO mature to support different decoding strategies of respective coarse temporal and sound intensity information at the brainstem level.
Lateral lemniscus¶
Inferior colliculus¶
Other¶
{Zoefel and VanRullen, 2016, #8293}
We presume that entrainment to low-level features of speech sound occurs relatively early in the auditory pathway (i.e., somewhere between cochlea and primary auditory cortex, including the latter; Davis and Johnsrude, 2003 and Lakatos et al., 2005), whereas entrainment to high-level features occurs beyond primary auditory cortex (Uppenkamp et al., 2006).
Summary¶
References¶
- Schnupp, J. W. H. (2013). Coding in the auditory system. In R. Q. Quiroga & S. Panzeri (Eds.), Principles of Neural Coding (pp. 179-204). CRC Press.
- Netter’s Atlas of Human Neuroscience. Author:David L. Felten, MD, PhD … Chapter:Sensory Systems Page:228; http://www.amazon.com/The-Netter-Collection-Medical-Illustrations/dp/1416063846
- Tzounopoulos, Thanos. “Mechanisms of Synaptic Plasticity in the Dorsal Cochlear Nucleus: Plasticity-Induced Changes That Could Underlie Tinnitus.” {American Journal of Audiology} 17.2 (2008): S170-75.
- Felix, Richard A., and Anna K. Magnusson. “Development of Excitatory Synaptic Transmission to the Superior Paraolivary and Lateral Superior Olivary Nuclei Optimizes Differential Decoding Strategies.” Neuroscience 334 (2016): 1-12.
Endnotes¶
| [1] | source of vertical axis |
| [2] | Author’s drawing, inspired on ‘Afferent Auditory Pathway’ in Netter (2013); see https://www.netterimages.com/afferent-auditory-pathways-unlabeled-physiology-frank-h-netter-1843.html |
| [3] | From https://commons.wikimedia.org/wiki/File:Afferent_%28PSF%29.png |
{kind=link}
Last edited Aug 26, 2019