Deciphering the Greek roots results in defining protozoa as 'first' (proto) 'animals' (zoa). Although molecular phylogenetic studies indicate that protozoa are among the earliest branching eukaryotes (see phylogenetic tree), such a definition does not provide much descriptive information. Protozoa are not easily defined because they are diverse and are often only distantly related to each other. Due to the extreme diversity of the protozoa the only feature common to all protozoa is that they are unicellular eukaryotic micro-organisms. Protozoa possess typical eukaryotic organelles and in general exhibit the typical features of other eukaryotic cells. For example, a membrane bound nucleus containing the chromosomes is found in all protozoan species. However, in many protozoan species some of the organelles may be absent, or are morphologically or functionally different from those found in other eukaryotes. In addition, many of the protozoa have organelles that are unique to a particular group of protozoa.
Questions one might ask in regards to defining protozoa
include:
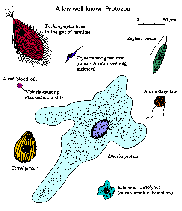
Protozoa exhibit a wide variety of morphologies (click figure for larger image and description). There is no one shape or morphology which would include a majority of the protozoa. Shapes range from the amorphous and ever-changing forms of ameba to relatively rigid forms dictated in part by highly ordered cytoskeletons or secreted walls or shells. Several protozoan species express photosynthetic or other pigments and thus are colored. Many protozoan species exhibit complex life cycles with multiple stages. Sometimes the different life cycle stages are so dissimilar that they have been mistaken for completely different species.
| Protozoan Sizes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
Protozoa are unicellular, or single-celled, organisms. However, some argue that they are actually 'acellular' since they are only composed of one cell and not multiple cells. Thus, the vast majority of protozoa are microscopic. However, they do exhibit an incredibly large range of sizes. Extant species range in size from <1 µm (10-6 meter) to several mm. Fossilized Forminiferida of several cm have been identified. (Extinct protozoa can be detected because of a secreted calcium carbonate shell.) Most of the organisms discussed in this course will be 3-50 µm. This small size necessitates the use of a microscope to detect protozoa. An electron microscope is needed for detailed morphological studies.
Protozoa are found in moist environments virtually everywhere. As a group, the protozoa are extremely adaptable. Individual species, though, generally have specific niches. Protozoa acquire and metabolize nutrients from their environment and thus are considered to be heterotrophic. Some protozoa simply absorb solutes (i.e., osmotrophy) from their media, while some are scavengers that ingest solid material (i.e., phagotrophy). Predatory protozoa either actively hunt down or passively ambush other organisms (typically bacteria or other protozoa). In contrast to protozoa, algae are autotrophic since they capture the energy of the sun and convert it to usable chemical energy (i.e., phototrophic). However, most unicellular algae are mixotrophic since they can combine heterotrophism and autotrophism.
Protozoa can also be viewed as free-living or symbiotic. Generally free-living organisms are found in the soil or aqueous environments, whereas symbionts live in close association with another organism. Symbiosis implies a physiological dependency of one organism on another organism and not just a close physical association between two organisms. Generally this dependency is in the form of nutrition. Different forms of symbiosis can be distinguished which reflect the nature of the association between the two organisms (Box).
| Symbiotic Associations | ||||||
|---|---|---|---|---|---|---|
|
 |
 |
 |
 |
| Modes of Motility | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
The earliest observations of protozoa noted their motility. This motility resulted in their classification as 'animals', which were distinguished from the non-motile 'plants'. However, motility is not a universal feature of protozoa and different protozoa utilize different mechanisms for their movement (Table). In fact, protozoa were initially classified based in part on their mechanism of motility (see Taxonomy).
Cilia and flagella are subcellular structures which propel protozoa through a fluid medium. Flagella are long whip-like structures which propel the organism as a result of wave-like beat which is propagated through their length. Flagellated protozoa typically have one or a few flagella per organism. In contrast, ciliated protozoa are usually covered with rows of numerous cilia. The beats of these cilia are coordinated and function like oars to propel the organism. Cilia and flagella can also assist in the procurement of food, reproduction and other functions. Cilia and flagella are made up of the same protein components and are actually equivalent structures. Both are membrane bound filamentous projections from the cell. The filament, known as an axoneme, is composed of a series of parallel microtubules, typically exhibiting a '9 + 2' arrangement. Movement is produced when the microtubules slide past each other. The force which mediates this sliding motion is generated by a protein called dynein. Dyneins are 'motor proteins' which convert the chemical energy released by ATP hydrolysis into a mechanical energy. Microtubules are cytoskeletal elements which also play important roles in cell shape and are a major component of the mitotic spindle.
In contrast to the swimming exhibited by flagellates and ciliates, ameba are protozoa that crawl along a solid substratum in a fashion known as 'ameboid movement'. The ameba projects out a pseudopodium, or false foot, from the cell body. The pseudopodium then attaches to the substratum and then pulls the rest of the cell body forward. The force involved in this movement is generated by another cytoskeletal system, which is comprised of actin and myosin. Actin forms long filaments, also known as microfilaments, and myosin is a motor protein which moves along the microfilaments in an ATP dependent manner. Muscle contraction is another example of the force generation via actin-myosin cytoskeletal elements. In a mechanistic sense, phagocytosis is a form of ameboid movement also involving microfilaments. In this case the pseudopodia are extended to surround the particle being ingested. Fusion of the pseudopodia with the cell body results in the internalization of the particle within a vacuole.
Apicomplexa also crawl along a substratum, but by a different mechanism than the ameba. The mechanism of this so-called 'gliding motility' is just beginning to be understood and probably involves both microfilament and microtubule based cytoskeletal systems. Apicomplexa also exhibit intracellular forms and invasion of the host cell also involves this gliding motility. (See also discussion of host cell invasion by the malaria parasite.)
Cellular motility involves force generation through either the microtubule-based cytoskeletal elements or the microfilament-based cytoskeletal elements. This is true for protozoa and other eukaryotes. The involvement of microtubules and microfilaments in both cell shape and cell movement make these subcellular structures more analogous to the musculoskeletal system.
|
|
|
|
| Modes of Reproduction | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Protozoa, like all other organisms, reproduce. The most common form of reproduction in protozoa is asexual binary fission. In other words, a single organism will divide into two equal organisms. A slight modification of this binary fission, called budding, is when one of the newly formed cells is smaller than the other. Typically the larger cell is called the mother and the smaller is the daughter. Some protozoa will form an intracellular bud and essentially give birth. Another variation of binary fission is a multiple fission or segmentation. In this situation, several rounds of nuclear replication occur without cytokinesis. This multinucleated cell will then form multiple progeny simultaneously.
Many protozoa exhibit sexual reproduction in addition to the asexual forms of reproduction. This sexual reproduction can involve the production and fusion of gametes in processes similar to higher organisms. The Ciliophora undergo a conjugation in which opposite mating types will pair and directly exchange genetic material (i.e., DNA). Sometimes sexual reproduction is an obligatory step in the life cycle, whereas in other cases the organism can reproduce asexually with an occasional round of sexual reproduction.
In summary, protozoa are unicellular eukaryotic microorganisms. However, the amount of diversity in terms of morphology, size and life styles exhibit by protozoa makes it difficult to develop a more precise definition. Their long evolutionary history (see phylogenetic tree) accounts for much of this diversity. However, protozoa do exhibit features common to all eukaryotes. (Link to a series of powerpoint presentations on eukaryotic cell biology.)
| Origin of Eukaryotic Cells |
|---|
The evolutionary jump from prokaryotes to eukaryotes is substantial and there are an estimated 5000 genes found in eukaryotes that are not found in prokaryotes (1). The first eukaryotic organisms likely resembled what we would now classify as protozoa. However, probably no direct descendants of the early eukaryotes currently exist and the path from prokaryotes to eukaryotes is likely littered with several missing links. Two key events in the evolution of eukaryotes are probably the development of internal cytoskeletal system and phagotrophy. A major difference between eukaryotes and prokaryotes is the rigid murein, or peptidoglycan, cell wall of bacteria. Although, many eukaryotes exhibit cell walls, they are fundamentally different than those of bacteria and are likely derived secondarily after the formation of the eukaryotes. For example, cell walls of plant cells are composed primarily of cellulose and fungal cell walls and cyst walls in many protozoa are composed primarily of chitin. The rigidity and strength provided by cell walls protects cells against mechanical stress and confers osmotic stability. Thus, coincident with the loss of the murein cell wall in the early eukaryote was the development of endocytoskeletal systems based on actin and tubulin to compensate for the relative fragility of a naked plasma membrane. In addition, the early eukaryotes likely evolved in a benthic environment and in particular in dense marine microbial mats. The organic solutes excreted by the bacteria and the high salt concentration of such an environment would have had an osmoprotective effect. The dense microbial mats would also provide an environment in which prey was very abundant and phagocytosis could have evolved without efficient cellular motility. The ancestral eukaryote probably attached to prey via cell surface N-linked glycoproteins that evolved after the loss of the murein cell wall and then digested them externally by secreting hydrolytic enzymes. To make the process more efficient the primitive predators could have progressively surrounded their prey until the prey were entirely engulfed in host membranes with digestion being carried out internally within phagosomes. This endocytic mechanism would also require the development of an exocytic mechanism to maintain the surface area of the plasma membrane. Thus, the evolution of phagocytosis not only included an expansion of the function of the actin cytoskeleton but also drove the development of membrane trafficking and subcellular compartments including the nucleus. The evolution of phagotrophy and endomembranes not only opened up a new way of eating, but also allowed for endosymbiosis and the enslavement of mitochondria. An early eukaryote probably engulfed an alpha-proteobacteria and formed a symbiotic relationship which evolved into an assimilation of the aerobic metabolism of this organism and the development of the mitochondrion. This enslavement of the mitochondrion was probably another key event in the evolution of the eukaryotes since extant eukaryotes are derived from mitochondria-containing ancestors. Extant anaerobic eukaryotes without mitochondria either have organelles derived from mitochondria, such as mitosomes and hydrogensomes, or at least have nuclear genes encoding remnant mitochondrial genes. Presumably the amitochondrial eukaryotic ancestor was at a competitive disadvantage and did not survive in the long term. Later a cyanbacterium was enslaved by the progenitor of plants and developed into the chloroplasts and similarly these plant predecessors were engulfed and enslaved to form the plastids found in many other algae and protozoa. 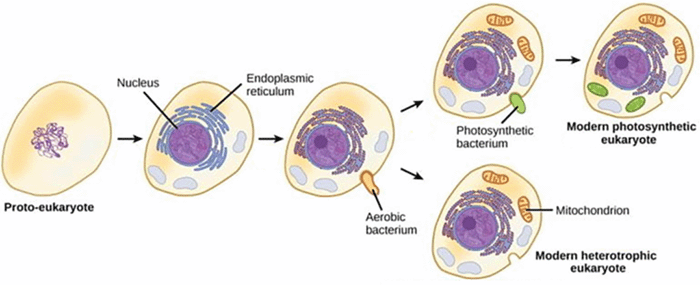
The cytoskeleton continued to evolve and expand its functions and in particular began to play a role in cellular motility. Most notably tubulin became a key component in flagella and later myosin evolved to allow for the development of ameboid movement and gliding motility. This motility allowed these early eukaryotes to expand into other niches (eg., planktonic, terrestrial) and thus this aerobic, phagotrophic and flagellated ancestral eukaryote could radiate into the diverse array of protozoa, and ultimately the other eukaryotes, that currently exist.
|
|
|
|
|
Taxonomy, or systematics, is the science of naming and classifying organisms. In addition to assigning hierarchical taxonomic classifications, systematics also attempts to place organisms into groups reflect evolutionary relationships or phylogenies. However, taxonomic criteria are often arbitrary and taxonomy is always changing to reflect new discoveries and interpretations. Furthermore, utilitarian features, such as type of disease, host range and geographical distribution, are frequently used in the systematics of pathogenic micro-organisms. This is sometimes called convenience grouping. Phylogeny is the representation of the evolutionary history and relationships between groups of organisms. Ideally taxonomic groups should be monophyletic groups in which all members of a group are derived from a common ancestor. However, convenience grouping or incomplete knowledge about the evolutionary histories often results in paraphyletic groups or polyphyletic groups.
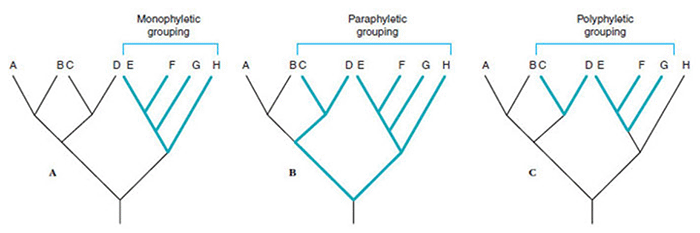
For example, many protozoa are closely related to species that are considered algae since there are phototrophic. Some have proposed to place unicellular eukaryotes, which includes protozoa, some algae, and unicellular fungi, into a single group called protista. This would make the protists a paraphyletic group including all eukaryotes except for multicellular organisms. Likewise, the protozoa are a paraphyletic group consisting of eukaryotes except multicellular organisms, algae, and fungi. The algae are a polyphyletic group consisting of phototrophic organisms ranging from prokaryotes to complex multicellular seaweed. The fungi are a monophyletic group consisting of unicellular and multicellular organisms.
Historically protozoa were divided into four major groups: the ameba, the flagellates, the ciliates, and the sporozoa. The distinguishing features between the groups was based on motility (i.e., ameboid, flagella, cilia). The sporozoa were a heterogeneous group that produced spores during one stage of their life cycles and exhibited a 'gliding' motility. However, such a classification scheme is quite arbitrary and does not necessarily reflect true evolutionary relationships between organisms. One problem with using motility as a taxonomic criteria is that many protozoa utilize different types of motility during different stages of their life cycles. For example, Naegleria exists in an ameba form when food is plentiful and transforms into a flagellate when food is absent. In general, the ameba are a heterogeneous group and are all probably dervived from flagellates. Among these four original protozoan groups only the ciliates are still considered a valid taxon.
Beginning the 1960's the electron microscope was used to identify ultrastructural features which could serve as criteria for grouping protozoa. In many cases morphology leads to a classification which places organisims into monophyletic groups. Monophyletic means that all of the organisms in that group are probably derived from a common ancestor. For example, many of the protozoa formerly called sporozoa possess subcellular structures, collectively known as the apical organelles, and now form a monophyletic group called apicomplexa. However, subcellular structures and metabolic pathways can be lost in some lineages and placing those descendants can be problamatic.
During the 1980's and continuing until present time molecular techniques are being applied to taxonomy. Possible evolutionary histories and relationships can be derived by comparing DNA or protein sequences. Molecular sequence data has confirmed phylogenies based on other criteria, settled some debates, and led to a few surprises. For example, molecular data confirms that the apicomplexa are monophyletic, and furthermore, indicates that they are related to the ciliates and dinoflagellates. These three groups are now combined into a larger monophyletic group called alveolata. This relationship had been previously suspected and the name is in reference to morphological structures known as alveolar sacs. But the use of single genes can be an unreliable means of determining evolutionary relationships, especially among distantly related organisms, and molecular data should be interpreted with caution.
Since the mid-1980’s classification of protists has been in a state of flux and afflicted with some philosophical controversies. On one hand, there is some argument for retaining elements of the Bütschlian scheme based on motility due to its familiarity and simplicity. However, these schemes are often in conflict with phylogenetic data. In addition, the hierarchies and ranks of the traditional Linnaean systematics (i.e., phylum, class, order, family, genus, species) does not always fit well with micro-organisms, and it is often difficult to decide which hierarchial level is most appropriate for any particular protozoan group. Furthermore, heterotrophic unicellular organisms (i.e., protozoa) are found in all of the major eukaryote groups.The last eukaryotic common ancestor (LECA) probably looked like and amoeboflagellate.
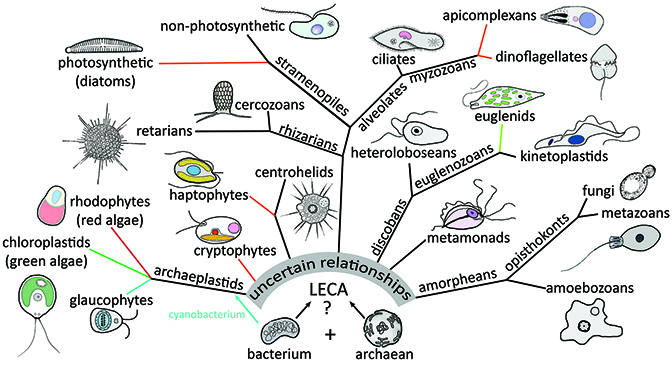
From: Chapter 64. Protozoa. In Encyclopedia of Biodiversity 3rd edition.